Larval Ecology
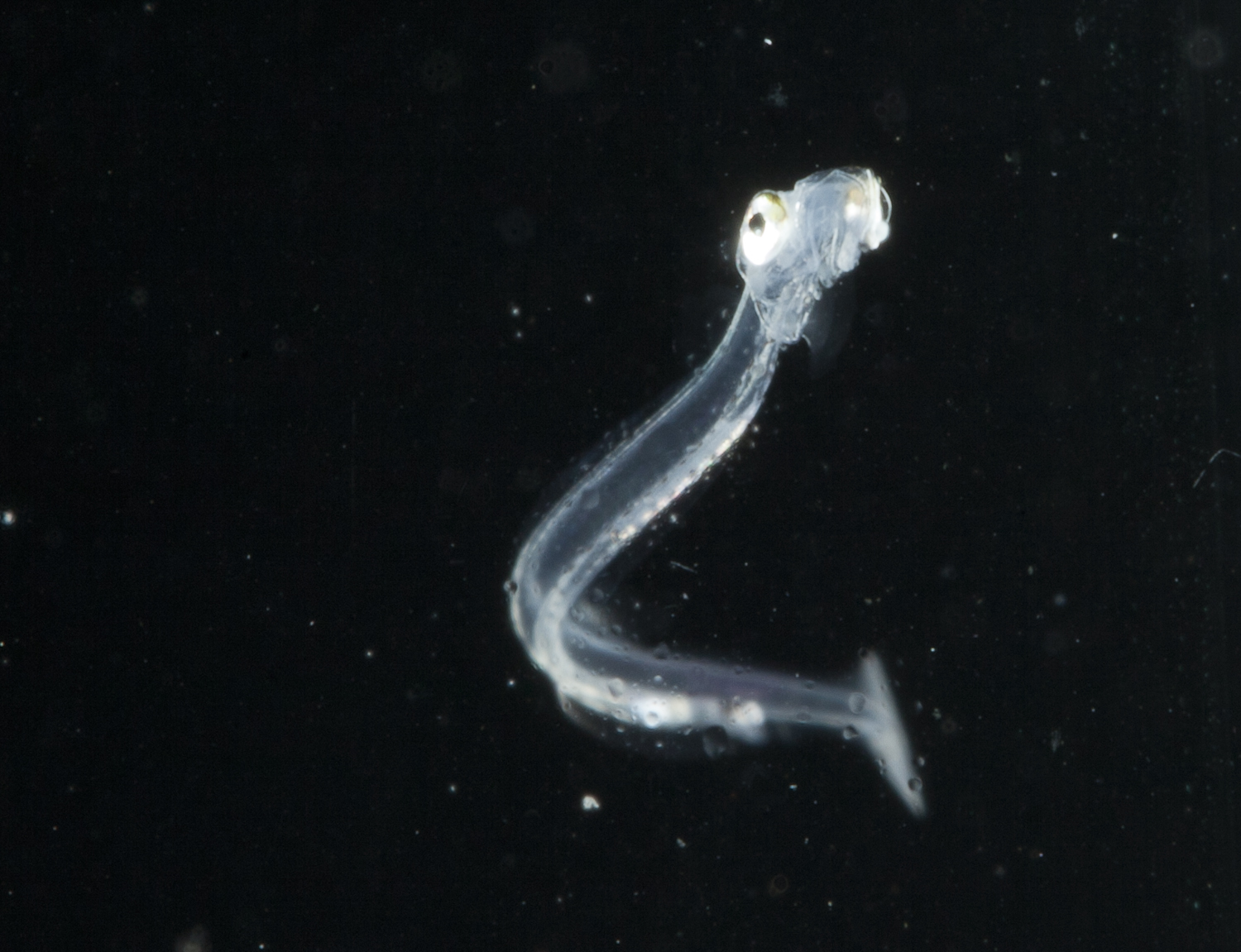
Survival of fish eggs and larvae is crucial for the recruitment to fisheries and the bottleneck in production for aquaculture. The focus of the larval ecology group therefore is on the effects of ocean currents, temperature, salinity and pH (ocean acidification) on development, growth, performance and survival of commercially important fish eggs and larvae, namely sprat, herring and cod. By monitoring the environment experienced by the larvae using field studies in the Baltic Sea (Bornholm Basin) and the Kiel Fjord and simulating changes in the environment as predicted by climate forecast scenarios in the laboratory, effects on fish eggs and larvae to be expected under future climate change scenarios can be studied. Additionally biological parameters like food availability, food quality and food web interactions (Dieckmann et al. 2012, Paulsen et al. submitted) as well as parental influences on larvae are a focus of the larval ecology group. Response variables measured in the larvae are biochemical indicators (essential fatty acid profiles, growth rates, RNA/DNA ratios and performance). Larval fish otoliths (the tinniest diary in the world) are analyzed for their increment structure in response to environmental stressors (Hinrichsen et al. 2010, Maneja et al. 2013). These indicators were calibrated (Meyer et al. 2012), evaluated under field settings and provided field evidence for starvation-induced recruitment regulation during the larval stage of cod in the Baltic Sea (Huwer et al. 2011). Analysing climate change scenarios on the different developmental stages of sprat using a stage specific Leslie- Matrix population growth model showed that temperature had the greatest impact on the growth rates of the feeding larvae confirming the importance of the larval stages for recruitment success (Haslob et al. 2012).

{kind=link}
{kind=link}
Effects of ocean acidification were studied on Baltic herring and cod larvae under laboratory conditions and indicated that the gametes and the early life stages were robust to pCO2 levels of up to 4000 ppm with no effect on growth and development accept for their protein biosynthesis capacity (Frommel et al. 2010, Franke & Clemmesen 2011, Frommel et al. 2012). Studies in large semi-natural outdoor mesocosms on cod and herring larvae of the Atlantic cod showed severe damages to important organs like the liver, kidney, pancreas, eye and gut, disturbances of the lipid metabolism as well as increase in otolith size under medium and high CO2 treatments giving first evidence of negative impact of ocean acidification on commercially important fish species from temperate waters (Frommel et al. 2012, Maneja et al. 2013). The results highlighted the importance of comparing fish stocks from different geographical regions, with differences in the variability of the environmental parameters. A direct comparison of Baltic cod and Canadian cod eggs and larvae showed significant effects of increased CO2 on fertilization and hatching success as well as genetic variability in response to climate stressors giving first evidence of an adaptation potential. References:Diekmann ABS, Clemmesen C, St. John MA, Paulsen, M, Peck, M (2012) Environmental cues and constraints affecting the seasonality of dominant calanoid copepods in brackish, coastal waters: A case study of Acartia, Temora and Eurytemora species in the Southwest Baltic. Mar. Biol. 159:2399–2414.Franke, A. and Clemmesen, C. 2011. Effect of ocean acidification on early life stages of Atlantic herring (Clupea harengus L.) Biogeosciences Discuss., 8, 7097–7126Frommel, A., Stiebens, V., Clemmesen, C. und Havenhand, J. (2010) Effect of ocean acidification on marine fish sperm (Baltic cod: Gadus morhua) Biogeosciences Discussions, 7 (4). pp. 5859-5872. ISSN 1810-6285Frommel, AY, Maneja, R, Lowe, D, Malzahn, M, Geffen, AJ, Folkvord, A, Piatkowski, U, Reusch, TBH, Clemmesen, C (2012) Severe tissue damage in Atlantic cod larvae under increasing ocean acidification. Nature Climate Change 2, 42-46.Frommel, A.Y., Schubert, A., Piatkowski, U., Clemmesen, C .(2012) Egg and early larval stages of Baltic cod, Gadus morhua are robust to high levels of ocean acidification. Mar Biol DOI 10.1007/s00227-011-1876-3Haslob, H., Hauss, H., Petereit, C., Clemmesen, C., Kraus, G., Peck, M.A. (2012) Temperature effects on vital rates of different life stages and implications for population growth of Baltic sprat. Mar. Biol. 159 . 2621-2632. Hinrichsen, H.H., Voss, R., Huwer, B., Clemmesen, C.: 2010: Variability of larval Baltic sprat (Sprattus sprattus L.) otolith growth: a modeling approach combining spatially and temporally resolved biotic and abiotic environmental key variables. Fish. Oceanogr. 19:6, 463-479Maneja RH, Frommel AY, Geffen AJ, Folkvord A, Piatkowski U, Chang MY, Clemmesen C. 2013: Effects of ocean acidification on the calcification of otoliths of larval Atlantic cod Gadus morhua. Mar. Ecol. Prog. Ser. 477:251-258.Meyer, S, Caldarone, EM, Chicharo, MA, Clemmesen, C, Faria, AM, Faulk, C, Folkvord, A, Holt, GJ, Hoie, H, Kanstinger, P, Malzahn, A, Moran, D, Petereit, C, Stottrup, JG, Peck, MA (2011). On the edge of death: Rates of decline and lower thresholds of biochemical condition in food-deprived fish larvae and juveniles. J. Mar. Syst. 93, 11-24. Paulsen, M, Clemmesen, C, Malzahn, AM. Essential fatty acid (docosahexaeonic acid, DHA) availability affects growth of larval herring in the field. In review at Mar. Biol.
back